BIOLOGÍA: EL ORÍGEN DE LAS ESPECIES. El origen de las especies (título original en inglés: On the Origin of Species) es un libro de Charles Darwin publicado el 24 de noviembre de 1859, considerado uno de los trabajos precursores de la literatura científica y el fundamento de la teoría de la biología evolutiva.
El origen de las especies
| El origen de las especies | |
|---|---|
| de Charles Darwin | |
 Página interior con el título de la edición de 1859. | |
| Género | Ensayo |
| Subgénero | Literatura científica |
| Tema(s) | Biología, biología evolutiva |
| Edición original en inglés (1859) | |
| Título original | On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life |
| Editorial | John Murray |
| Ubicación | |
| Edición traducida al español (1877) | |
| Traducción | Enrique Godínez y Esteban y Antonio Zulueta |
| Editorial | Biblioteca Perojo |
| Ubicación | |
| ISBN | 978-84-206-6867-3 |
El origen de las especies (título original en inglés: On the Origin of Species) es un libro de Charles Darwin publicado el 24 de noviembre de 1859, considerado uno de los trabajos precursores de la literatura científica y el fundamento de la teoría de la biología evolutiva.
El título completo de la primera edición fue On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (El origen de las especies por medio de la selección natural, o la preservación de las razas favorecidas en la lucha por la vida). En su sexta edición de 1872, el título corto fue modificado a The Origin of Species (El origen de las especies).
En esta obra Charles Darwin (1809-1882) propuso el mecanismo de la selección natural como explicación para el origen de las especies. Haciendo un paralelismo con la selección artificial realizada por el ser humano en plantas o ganado, Darwin creó el concepto de selección natural, en el cual la naturaleza selecciona a las poblaciones más aptas para la supervivencia en determinado ambiente y descarta a las menos aptas.
En las primeras ediciones, Darwin no empleó el término «evolution» («evolución»), sino expresiones como «descendencia común con modificación» o similares; solo desde la 5ª, y así aparece en la 6ª de 1876, la última corregida y revisada por él mismo, aparece el término «evolución». Por lo demás, en ningún momento centra su explicación en la especie humana.[1]
Contenido[ocultar] |
[editar] Elaboración
Los hechos expuestos en El origen de las especies fueron acumulados por Darwin mismo a lo largo de su viaje en el HMS Beagle entre 1831-1836. Sin embargo, no fue hasta la lectura del ensayo de Thomas Malthus sobre el principio de la población que Darwin dio con un marco teórico que consideró adecuado para hilar la argumentación de su obra:
El libro se puso a la venta el 24 de noviembre de 1859, en la editorial John Murray de Londres, y agotó los 1.250 ejemplares impresos en el primer día.
[editar] Publicación

Como se pone de manifiesto en El origen del hombre, y la selección en relación al sexo (The Descent of Man, and Selection in Relation to Sex), Darwin había reflexionado ampliamente en las implicaciones de su teoría sobre el origen de la humanidad, si bien el tema de la evolución humana no había sido tratado en profundidad en "El Origen de las especies".
La publicación de sus ideas sobre la evolución fue adelantada a causa de la investigación independiente de una teoría similar realizada por Alfred Russel Wallace en 1858 -el libro de Darwin fue publicado en 1859-. Muchos consideran que Wallace merece tanto crédito como Darwin por su teoría de la selección natural, aunque la obra de Darwin presenta su teoría con una mayor cantidad de observaciones y una mejor argumentación.
Charles Darwin hizo así mismo muchas de sus investigaciones, llegando así a sus propias teorías, con métodos rudimentarios y puramente caseros. En el jardín de su casa observaba con instrumentos muy básicos la evolución de las plantas, y así de forma completamente empírica y poco sofisticada llegaba a sus conclusiones.
En relación con la publicación de El origen de las especies en español puede consultarse la sección Obra de Charles Darwin en español del artículo Obra de Charles Darwin.
[editar] Contenido
Ernst Mayr distingue cinco subteorías en el Origen: el hecho de la evolución, la postulación de un origen común para todos los organismos, la diversificación de las especies, el gradualismo y la selección natural.[3] [4] Michael Ruse distingue entre el hecho de la evolución, el patrón evolutivo (curso real del concreto proceso histórico ocurrido desde el origen de la vida hasta la actualidad) y la teoría de la evolución (explicación teórica del cambio).[5]
[editar] La comunidad de descendencia
Mediante la teoría del origen común, Darwin logró integrar armoniosamente evidencias procedentes de campos tan dispares como la biogeografía, la paleontología, la anatomía comparada o la embriología. La convergencia de todas estas evidencias demostraba la comunidad de descendencia de todos los organismos vivos y extintos. De este modo, Darwin ofrecía una demostración sistemática del transformismo, oponiéndose al fijismo (defendido en el marco tanto del uniformismo como del catastrofismo) y a la teoría de las creaciones sucesivas:
[editar] El origen de las variaciones
Darwin admite un abanico muy amplio de causas de variabilidad:
- Las condiciones de vida: según Darwin, las condiciones de vida pueden ejercer una acción directa (cuando actúan sobre todo el organismo o sobre ciertas partes) o indirecta (sobre el aparato reproductor). En el primer caso, los efectos en la descendencia pueden ser determinados o indeterminados: son determinadas las modificaciones que afectan a la totalidad (o a la práctica totalidad) de los individuos de una misma especie, dada su exposición durante varias generaciones a ciertas condiciones ambientales; son indeterminadas las pequeñas particularidades que distinguen a los individuos de una misma especie como resultado de la exposición de cada organismo a las condiciones de vida y que no pueden explicarse por herencia.
- El uso y del desuso: en el Origen, Darwin admite también el efecto lamarckiano del uso y desuso de los órganos (pp. 200-201)1. El problema no es, por tanto, el de la incompatibilidad causal, sino el de discernir, en cada caso, las transformaciones debidas a la selección natural, al uso y al desuso o a su combinación (pp. 208-210).
- La variación correlativa: el término “variación correlativa” comprende, en realidad, tres tipos de variabilidad: la variación entre los cambios ocurridos en el embrión y su traducción en el animal adulto, la ley de la compensación y economía del crecimiento y la variación correlativa entre órganos. Algunas son admitidas completa o parcialmente; otras quedan integradas en la selección natural.
[editar] La probabilidad de la aparición de variedades
La selección natural no crea las variaciones individuales, sino que las utiliza como material de construcción, como el hombre para crear variedades domésticas (p. 95). Lo único que puede hacer la selección natural es conservar y acumular variaciones útiles. “Si no aparecen éstas, la selección natural no puede hacer nada” (p. 132). Pero ¿cuáles son las circunstancias que influyen en la producción de variabilidad? Darwin ofrece varias causas al respecto:
- La variabilidad puede variar entre los individuos, y el índice de variabilidad es heredable (p.178).
- La producción de variabilidad depende del número de individuos sobre los que actúa la selección: cuanto mayor sea, mayor probabilidad de que surjan variaciones favorables. De ahí que las especies que pertenecen a géneros mayores sean las que con más frecuencia presentan variedades. Puesto que la selección natural obra mediante formas que tienen alguna ventaja sobre otras en la lucha por la existencia, actuará principalmente sobre aquellas que tienen ya una ventaja, y la magnitud de un grupo muestra que sus especies han heredado de un antepasado común alguna ventaja en común. Por consiguiente, la lucha por la producción de descendientes nuevos y modificados será principalmente entre los grupos mayores, que están todos esforzándose por aumentar en número. Un grupo grande vencerá lentamente a otro grupo grande, lo reducirá en número y hará disminuir así sus posibilidades de ulterior variación y perfeccionamiento. Dentro del mismo grupo grande, los subgrupos más recientes y más perfeccionados, por haberse separado y apoderado de muchos puestos nuevos en la economía de la naturaleza, tenderán constantemente a suplantar y destruir a los subgrupos más primitivos y menos perfeccionados. Los grupos y subgrupos pequeños y fragmentarios desaparecerán finalmente. (p. 186) La subordinación de unos grupos a otros queda explicada por la tesis de que las especies con mayor variabilidad son las de mayor distribución. Así, los grupos grandes tienden a continuar aumentando. Y como los descendientes que varían de cada especie procuran ocupar el mayor y más diferente número de puestos posibles, tienden constantemente a divergir en sus caracteres. Por último, las formas que aumentan en número y divergen en caracteres tienen una tendencia a suplantar y exterminar a las formas precedentes menos divergentes y perfeccionadas.
- De este modo se explican dos hechos siempre presentes en las clasificaciones: 1) “ todos los organismos vivientes y extintos están comprendidos en un corto número de grandes órdenes y en un número menor de clases.” (p. 572) y 2) “los descendientes modificados procedentes de un progenitor, quedan separados en grupos subordinados a otros grupos” (p. 553).
- El tiempo es también un factor determinante: a mayor tiempo, mayor probabilidad de que aparezcan variedades.
- Según Darwin, los cambios en las condiciones de vida producen una tendencia a aumentar la variabilidad (p.139).
- La existencia de “nichos vacíos” que puedan ser explotados sin competencia.
[editar] La selección natural
En el Origen, Darwin utilizó la selección artificial como una analogía fundamental para la comprensión del mecanismo de la selección natural. La analogía de las técnicas agrícolas y ganaderas había sido ya utilizada por Lamarck como evidencia de la eficacia de su ley de uso y desuso de los órganos (Lamarck, PhZ, p. 226). También Darwin, instigado por John Herschel, encuentra en la analogía un gran aliado metodológico. Tanto la selección artificial como la selección natural tienen como resultado la transformación de las especies gracias a la acumulación progresiva de variaciones. La gran diferencia estriba en la dirección del cambio: dirigida hacia la utilidad del hombre, en un caso, ciega en el otro (p. 81). Sin embargo, en muchos casos la selección artificial se remonta a épocas tan remotas, que su efecto resulta inconsciente para los hombres (p. 88).
La supervivencia del más fuerte “incluye no sólo la vida del individuo, sino también el éxito al dejar descendencia” (p. 118).
La influencia de la teoría de Malthus en la formulación de la teoría de la selección natural se reconoce explícitamente en El Origen:
La teoría de la selección natural logra explicar multitud de hechos biogeográficos:
[editar] Gradualismo
La ausencia o rareza de variedades de transición en el registro fósil fue una de las objeciones más repetidas a la teoría darwiniana. En el capítulo "Dificultades de la teoría", Darwin alegó distintas razones para explicar la ausencia de variedades intermedias:
- La transformación de partes aisladas en territorios actualmente continuos.
- Las variedades más numerosas tendrían mayor ventaja evolutiva y harían desaparecer a las minoritarias.
- La lucha entre las especies de un mismo género es más encarnizada (p. 182).
- Enfrentándose a Lyell, quien oponía la fragmentariedad del registro fósil al gradualismo filogenético, Darwin lo califica de incompleto. El capítulo "De la imperfección de los registros geológicos" está destinado a refutar los “hechos” que desde la paleontología se le objetaron al gradualismo de la teoría evolutiva.
[editar] Divergencia de caracteres
Según el principio de la divergencia, los grupos con más géneros resultaban ser los que presentaban más especies y más subespecies. Darwin lo explica a partir de la selección natural: los grupos biológicos obtienen ventajas al diferenciarse lo más posible, en forma similar a como las obtienen los miembros de un mismo grupo al diferir entre sí (Origen, p. 172). Darwin comparaba el principio de la divergencia con la “división fisiológica del trabajo” de Henri Milne-Edwards, que sostenía que mientras más especializadas son las distintas partes del organismo más eficaz es el organismo en su conjunto. (Origen, p. 242)
[editar] El concepto de especie
En el Origen, Darwin ofrece varios argumentos contra la concepción morfológica de especie. Así, recurre al dimorfismo sexual y otros polimorfismos (la alternancia de las generaciones, de larvas frente a los adultos y de las diferentes formas de flores que existen en una serie de especies de plantas) para demostrar que el concepto morfológico de especie no tiene ningún sentido como base adecuada para la construcción de un lenguaje biológico.[6] Sin embargo, el concepto de especie defendido por Darwin continúa siendo una cuestión controvertida. Según Mayr, sus cuadernos de notas muestran que hacia 1837 había abandonado el concepto tipológico de especie, desarrollando un concepto biológico basado en el aislamiento reproductivo. Sin embargo, argumenta Mayr, quince años más tarde, a partir de sus estudios de variedades de plantas, abandonó el concepto biológico para volver a una definición entre tipológica y nominalista como la defendida en el Origen.[7]
Ghiselin sostiene que el problema es más complicado y que el concepto darwiniano de especie se acerca considerablemente al concepto biológico de la síntesis evolutiva moderna. Aunque nunca llegó a defender la definición biológica de especie en su sentido estrictamente moderno, es decir, las especies como poblaciones reproductivamente aisladas, Ghiselin sostiene que Darwin consideraba a las especies como unidades evolutivas y, por lo tanto, reales. Basándose tanto en declaraciones explícitas de Darwin en los cuadernos de notas como en su práctica sistemática, Ghiselin demuestra que "Darwin no consideró que las especies fueran necesaria y totalmente arbitrarias y que no se basó simplemente en la distinción y la semejanza morfológica".[8] Lo que negaba Darwin no era la realidad de los taxones sino de las categorías taxonómicas.[9]
[editar] Véase también
[editar] Referencias
- ↑ Cf. Barbara Continenza, "Presentación" y "La evolución, una revolución", en Investigación y Ciencia. Temas 54. Darwin.
- ↑ Darwin, 1958, p. 120
- ↑ Mayr, E. (1992, pp.48-49). Una larga controversia: Darwin y el darwinismo. Santos Casado de Otaola (trad.). Crítica.
- ↑ Las subteorías referidas por Mayr no son todas las teorías evolutivas de Darwin, (selección sexual, pangéneis, efecto del uso y desuso y divergencia de caracteres) sino aquellas que se han tenido en cuenta a la hora de comentar la obra de Darwin.
- ↑ Ruse, M. (1983). La revolución darwinista:(la ciencia al rojo vivo). C. Castrodeza (trad.). Alianza.
- ↑ Ghiselin, 1983, p. 118
- ↑ Mayr, 1995, p. 43
- ↑ Ghiselin, 1983, p. 112
- ↑ Ghiselin, 1983, p. 113-114
[editar] Bibliografía
- Mayr, Ernst (2001). Una larga controversia: Darwin y el darwinismo. Casado de Otaola, Santos (trad.). Barcelona: Editorial Crítica. pp. 43. ISBN 978-84-8432-254-2.
- Ghiselin, Michael T. (1983). El triunfo de Darwin (1ª ed, edición). Cátedra. pp. 272 págs.. ISBN 9788437604305.
[editar] Enlaces externos
 Wikisource contiene una copia de El origen de las especies.Wikisource
Wikisource contiene una copia de El origen de las especies.Wikisource
- En Biblioteca de Traductores pueden verse y descargarse los 3 tomos: Tomo I Tomo II Tomo III de El origen de las especies, Espasa Calpe, 1ª edición, 1921, traducción al español por el biólgo Antonio Zulueta
- El origen de las especies. texto íntegro -Cervantes virtual-, traducido al español por el biólogo Antonio Zulueta
- Obra completa de Charles Darwin -en línea-, bibliografía de On the Origin of Species:
Ambas páginas con enlaces a textos e imágenes de todas las ediciones de The Origin of Species, incluyendo traducciones al alemán, danés y ruso. - Texto completo Origien of Species 1ª edición -en pdf- en la Universidad de Nuevo Gales del Sur (University of New South Wales)
- Portal de biología de la evolución




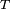 para un conjunto de lenguas
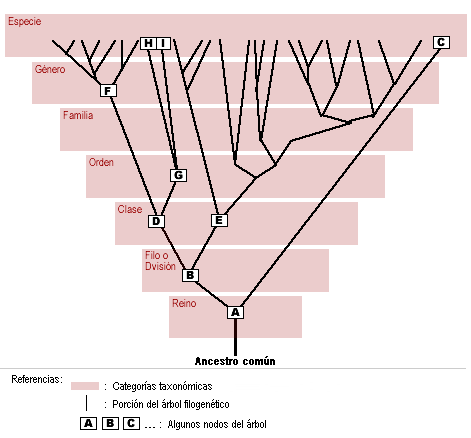
para un conjunto de lenguas 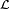 tiene filogenia perfecta si para un conjunto de caracteres
tiene filogenia perfecta si para un conjunto de caracteres 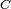 , si es posible etiquetar los nodos del árbol de tal manera que los caracteres evolucionen a lo largo del árbol sin retromutaciones o evolución paralela en
, si es posible etiquetar los nodos del árbol de tal manera que los caracteres evolucionen a lo largo del árbol sin retromutaciones o evolución paralela en 










.jpg)















{kind=link}